Физиологические аспекты
Спинномозговая жидкость
Спинномозговая жидкость (СЖ) является прозрачной бесцветной жидкостью, чьи физико-химические свойства показаны в таблице 2.1.
Таблица 2.1 Физико-химические свойства СМЖ
| Относительная плотность | 1.003-1.09 |
| Плотность | 1.0001-1.0005 (37°С) |
| Баричность | 1.000 |
| pH** | 7.31-7.34 |
| Глюкоза** | 50-75 мг/дл |
| Протеин** | 18-41 мг/дл |
| Общее число клеток** | 2-5 ?/л |
| Натрий** | 137-153 ммоль/л |
| Калий** | 2.6-3.3 ммоль/л |
| Кальций** | 1.02-1.34 ммоль/л |
| Магний** | 0.9-1.2 ммоль/л |
| Хлорид** | 120-130 ммоль/л |
| Давление* | 9-20 см водяного ст. |
| * Давление указано при положении лежа. ** Химические свойства взяты из таблиц Geidy Scientific Tables (1981) Vol 1, !65-177. |
|
Девяносто процентов СЖ формируется в сосудистых сплетениях (небольших красноватых пучках или участках тканей) в боковых, третьем и четвертом желудочках (рис.1.38), тогда как 10 процентов образуется в веществе мозга. Кровь и СЖ разделяются непроницаемыми “плотными соединениями” между эпителиальными клетками (рис.2.1).
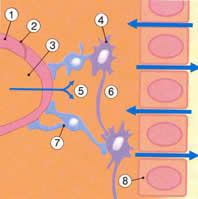
| Рис.2.1. Основные нутриенты достигают нейронов и глиальных клеток головного мозга, проходя или через гематоэнцефалический барьер (кровь-СМЖ), который регулируется сосудистым сплетением, или через барьер церебральных капилляров. Водорастворимые молекулы не могут свободно диффундировать между кровью и спинно-мозговой жидкостью из-за непроницаемых жестко связанных соединений между эпителиальными клетками сосудистых сплетений, вместо этого эпителиальные клетки переносят определенные молекулы с одной стороны барьера на другую. Как только молекулы попадают в СМЖ, они диффундируют через «протекающий» эпителиальный слой и достигают интерстициальной жидкости, окружающей нейроны и глиальные клетки. |
|
 |
 |
| Рис. 2.1а. 1.Эндотелиальная клетка 2.Плотное соединение 3.Церебральный капилляр 4.Нейрон 5.Глюкоза 6.Интерстициальная жидкость 7.Глиальная клетка 8.Эпендимный слой |
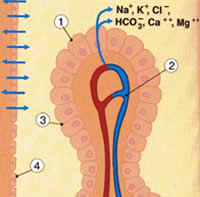
Рис.2.1б. 1.Хориоидальное сплетение, эпителиальная клетка 2.Капилляр 3.Плотное соединение 4.Эпендимный слой |
Эпителиальные клетки переносят определенные молекулы из капилляров внутрь желудочков головного мозга (рис.2.2). Молекулы воды в эпителиальных клетках диссоциируют на ионы водорода и гидроксильные ионы. Гидроксильные ионы комбинируются с двуокисью углерода, которая является продуктом клеточного метаболизма. На поверхности базолатеральных клеток ионы водорода обмениваются на внеклеточные ионы натрия из плазмы. В желудочках мозга ионы натрия активно переносятся через апикальную поверхность клетки. Это сопровождается компенсаторным движением ионов хлорида и бикарбоната в СЖ. Для поддержания осмотического равновесия вода движется в желудочки (рис.2.2).
Рис. 2.2. Поток ионов, пересекающий гематоэнцефалический барьер (кровь-СМЖ) регулируется несколькими механизмами в сосудистом сплетении. |
|
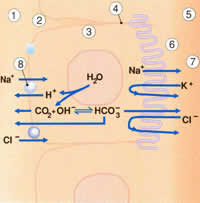 |
1.Кровеносный сосуд (плазма) 2.Базолатеральная поверхность 3.Эпителиальная клетка сосудистого сплетения 4.Жесткая связь 5.Желудочки 6.Апикальная поверхность 7.СМЖ в желудочке 8.Ионный обмен |
Объем СЖ, производимый ежедневно, предположительно равен объему, находящемуся в центральной нервной системе (ЦНС), который составляет приблизительно 150 мл. Однако, при потере СЖ (напр., во время или после люмбальной пункции), значительное количество ее может быть продуцировано, чтобы скомпенсировать потерю.
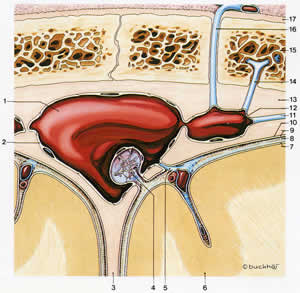
Скорость абсорбции СЖ зависит от давления и линейно в широком диапазоне. Арахноидальные грануляции, находящиеся в краниальных венозных синусах, дренируют основную часть СЖ. Когда давление СЖ превышает таковое в венозных синусах, тонкие канальцы в грануляциях позволяют СЖ стекать внутрь синусов. И наоборот, когда давление в венозных синусах превышает давление СМЖ, окончания грануляций закрываются, предотвращая обратный отток (рис.2.3). Другие возможные участки абсорбции включают вены в субарахноидальном пространстве и в лимфатических сосудах краниальных и спинальных нервов.
| Рис 2.3. | 1.Верхний сагиттальный синус 2.Эндотелий 3.Мозговой намет 4.Арахноидальная грануляция 5.Верхняя церебральная вена 6.Кора головного мозга 7.Мягкая мозговая оболочка 8.Субарахноидальное пространство и арахноидальная трабекула 9.Арахноидальная мембрана 10.Субдуральное пространство 11.Менингеальная вена 12.Полость вены 13. Твердая мозговая оболочка 14.Вена губчатого вещества (кости) 15.Пристеночная кость (cranium ) 16.Эмиссарная (дренирующая) вена 17. Pericranium |
 |
Помимо физической защиты головного мозга, СЖ обеспечивает ему и химически стабильную окружающую среду. Разделение крови и СЖ регулирует важнейшие биохимические функции в головном мозге. Кроме того, это разделение, вероятно, принимает участие в питании мозга и помогает удалять продукты метаболизма. Более того, оно является важным в распределении и удалении лекарственных средств, введенных в СЖ (напр., местных анестетиков и опиоидов).
Физиология сенсорных афферентных путей
Ноцицепторы и периферические механизмы боли
Ноцицепторы являются специфическими болевыми рецепторами, активируемыми только интенсивной, разрушающей ткань активностью (действием). Структурно ноцицепторы являются свободными нервными окончаниями, локализованными в коже и других чувствительных к боли тканях. А-дельта или С-волокна проводят афферентные импульсы в ответ на болевой стимул и классифицируются в соответствие со стимулом, который их активирует. (Табл.2.2).
Таблица 2.2. Ноцицепторы
| Тип рецептора | Стимул | Типы волокон | Скорость передачи (м/сек) |
| Механический | Механический | А - дельта | 2-30 (миелинизированные) |
| Механотермический | Механический и термический | А - дельта | 2-30 (миелинизированные) |
| Полимодальный | Механический, термический и химический | С - волокна | <2(немиелинизированные) |
Интенсивная механическая стимуляция активирует механические ноцицепторы . Механотермические ноцицепторы отвечают на повреждающую механическую и термическую стимуляцию. Полимодальные ноцицепторы отвечают на интенсивные механические, термические и химические стимулы (воздействия).
Ноцицептивные импульсы передаются по быстрым А-дельта волокнам (скорость передачи = 2-30 м/сек), которые достигают заднего рога спинного мозга через дорсальные спинномозговые нервные корешки.
Анатомия заднего рога
Задний рог анатомически разделен на шесть тонких слоев (нейрональные пластины I - VI по Rexed ) (рис.2.4).
| Рис.2.4. Сенсорные афферентные пути | |
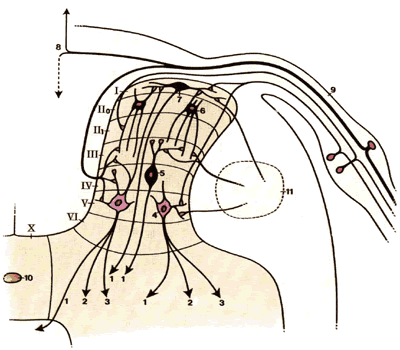 |
1.К контралатеральному клиновидному пучку 2.К переднему рогу 3.К ипсилатеральному клиновидному пучку 4.Нейроны широкого динамического действия 5.Клетка «антенны» 6.Клетка ножки 7.Маргинальная клетка 8.Дорсальная колонна 9.Дорсальный корешок 10.Центральный канал 11.Дорсолатеральный пучок (нисходящее ингибирование) |
Слой I (Маргинальная зона)
Маргинальная зона заднего рога состоит из больших противоположно ориентированных нейронов. Некоторые клетки слоя I направлены непосредственно в таламус через контралатеральные восходящие пути, тогда как другие восходят внутри- и/или межсегментарно через задние и заднебоковые пути. Нейронные клетки слоя I отвечают на:
- Сильное механическое воздействие (А-дельта волокна)
- Температуру более 45°С (А-дельта волокна)
- Повреждающее механическое, термическое или химическое воздействие (полимодальные С-волокна)
Слой II (Желатинозная субстанция)
Слой II разделен на внутреннюю ( IIi ) и внешнюю ( IIo ) области. Главной клеткой слоя II является стволовая клетка, чье дендритическое дерево разветвляется через слой II в слой III и чьи аксоны проецируются в слой I . Нейроны слоя II возбуждаются механическими или термическими ноцицептивными афферентами. Нейроны с дендритами в IIo активируются ноцицептивными волокнами, тогда как таковые с дендритами в слое IIi активируются безопасным (неповреждающим) механическим воздействием.
Слой III-V (Nucleus proprius или magnocellular area)
Дендриты относительно больших нейронов слоя IV распространяются поперечно (крест-накрест) и дорсально в желатинозную субстанцию. Клетки в слое IV отвечают на царапины и прикосновения, но при усилении этих воздействий не увеличивают свою активность. Нейроны широкого динамического диапазона (ШДД) слоя V являются мультирецептивными нейронами, которые отвечают на неповреждающее прикосновение, низкие или высокие механические, термические и химические стимулы (воздействия). Активность ШДД-нейронов возрастает с ростом интенсивности раздражения.
Нейроны слоя V являются важным звеном спинно-таламического, спинно-ретикулярного и спинно-мезэнцефалического путей (см. ниже), поскольку вносят существенный вклад в их функционирование.
Слой Х (Центральный канал)
Слой Х не является частью дорсального рога. Однако, клетки в этой области имеют свойства, такие же как и маргинальные клетки слоя I и активируются воздействием высокой или низкой температуры и повреждающими механическими раздражениями, участвуя в процессе формирования и передачи боли.
Механизмы дорсального рога
Клеточные пересечения в дорсальном роге являются основными в механизме обработки болевого сигнала. Однако, механизмы, ответственные за передачу боли, невероятно сложны. Здесь может быть представлен лишь их краткий обзор.
Когда нервные волокна входят в спинной мозг, они разделяются во входной зоне дорсального корешка таким образом, что большие волокна локализуются медиально, а меньшие волокна – латерально. Волокна ноцицепторов проходят прямо во внешние слои серого вещества заднего рога, где они образуют синапсы с маргинальными клетками слоя I , стволовыми клетками слоя II , ШДД-нейронами слоя V и клетками серой спайки (слой Х).
А-бета волокна, проводя безболевую тактильную или прориоцептивную информацию, входят в спинной мозг медиально по отношению к дорсальному рогу, где они раздваиваются и посылают ветви краниально в дорсальные колонны и в клетки в слоях II - V (рис.2.4).
Клетки в слое I отвечают почти исключительно на повреждающие стимулы. Аксоны этих клеток заканчиваются в более глубоких слоях в дорсальном роге или нисходят к контралатеральному таламусу ( vide infra ). К ШДД-нейронам в слое V подходят как большие, так и малые волокна, многие из которых заканчиваются в контралатеральном ядре таламуса. Ингибиторные интернейроны, локализованные в желатинозной субстанции, широко соединены с другими нейронами дорсального рога, которые также нисходят в таламус.
Эти ингибиторные клетки модифицируют активность ноцицептивных нейронов. Сегментарные и нисходящие неврологические пути ( vide infra ) влияют на ингибиторную активность клеток желатинозной субстанции (ЖС) (рис.2.4).
Неврологическая передача в спинном мозге
Болевые стимулы приводят к высвобождению нейромедиаторов из ноцицептивных нейронов, которые активируют нейроны второго порядка, заканчивающиеся в верхних центрах. Интернейроны в ЖС могут модулировать высвобождение этих нейромедиаторов, активируя ингибиторные пресинаптические рецепторы. Считается, что этими ингибиторными пресинаптическими нейромедиаторами являются энкефалины. Интраспинальные опиоды вызывают сегментарную анальгезию предположительно путем активации тех же пресинаптических ингибиторных рецепторов, что и энкефалины, блокируя, таким образом, высвобождение болевых нейротрансмиттеров (Рис.2.5).
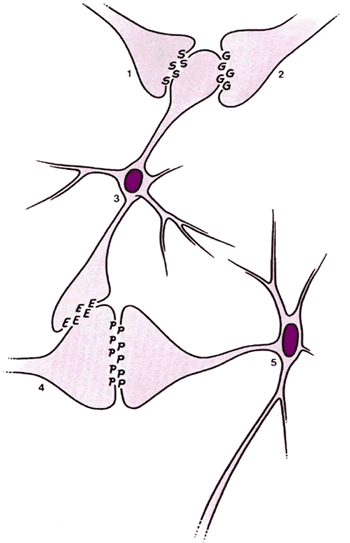 |
Рис.2.5 Предложенный механизм ингибирования активации болевых нейронов посредством энкефалин-секреторных интернейронов. Болевые нейроны активируются высвобождением нейротрансмиттерного вещества из ноцицепторов. Ингибиторные интернейроны влияют на тоническое пресимпатическое ингибирование высвобождения болевых нейротрансмиттеров. Ингибирование увеличивается сегментарным, а также нисходящим входом. 1.Нисходящий ингибиторный нейрон из области околоводопроводного серого вещества. 2.Большой А-бета нейрон 3.Энкефалинэргический интернейрон в желатинозной субстанции 4.Ноцицептор (А-дельта иди С-волокно) 5.Нейрон второго порядка, ведущий к спинно-таламическому пути G = гамма-аминобутиратная кислота S = 5-гидрокситриптами или норэпинефрин E = энкефалин P = нейротрансмиттер боли («Субстанция Р») |
5-гидрокситриптамин (5НТ, серотонин) и норэпинефрин также модулируют высвобождение болевых нейротрансмиттеров в ЖС путем активации нисходящих ингибиторных путей ( vide infra ). Неврологическая активность в больших А-бета волокнах в том же сегменте может также подавлять ответ на повреждающие стимулы в ЖС (так называемое закрытие ворот), которые, как полагают, обусловлены высвобождением ГАМК (гамма-аминомасляной кислоты). Это взаимодействие больших и малых волокон в ЖС является базовым элементом теории воротного контроля, предложенной Мелзаком и Уоллом. Были предложены и другие возбуждающие/ингибирующие медиаторы, но их описание выходит за рамки этой публикации.
Восходящие болевые пути
Спинно-таламический тракт (СТТ)
Восходящие болевые пути, которые заканчиваются в вентробазальной и задней частях таламуса, берут начало в маргинальных клетках слоя I и ШДД-нейронах слоя V . Хотя несколько аксонов восходят ипсилатерально, большинство пересекают вентрально белое вещество по направлению к противоположному вентролатеральному канатику и направляются рострально, формируя латеральный спинно-таламический тракт (СТТ) (рис.2.6). Некоторые волокна СТТ заканчиваются в вентральном постлатеральном ядре, тогда как другие заканчиваются в медиальной части заднего таламуса. Эта часть СТТ касается пространственных и временных аспектов болевой рецепции и известна как неоспинно-таламический тракт. Волокна неоспинно-таламического тракта после образования синапса в таламусе проецируются на кору головного мозга. Другие волокна СТТ, которые заканчиваются в ретикулярной формации, варолиевом мосте, среднем мозге, сером околоводопроводном веществе (СОВ), гипоталамусе, медиальном и интраламинарном таламических ядрах, образуют палеоспинно-таламический тракт и касаются аспектов отмены и вегетативных ответов на боль.
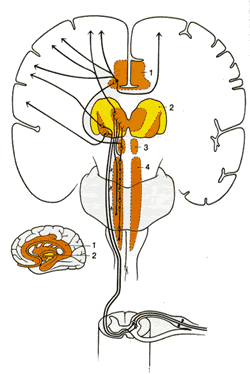 |
Рис.2.6. Диаграмма показывает пути вдоль латерального спинно-таламического тракта из спинного в головной мозг. 1.Лимбическая структура переднего мозга 2.Таламус 3.Гипоталамус 4.Ретикулярная формация |
Спинно-ретикулярный тракт (СРТ)
СРТ берет начало в слоях I , V , VII , VIII и Х, и восходит билатерально, заканчиваясь в ядрах ретикулярной формации головного мозга (рис.2.7). Эти ядра модулируют ноцицептивную информацию и большей частью касаются раздражающих, вегетативных и мотивационных аспектов боли.
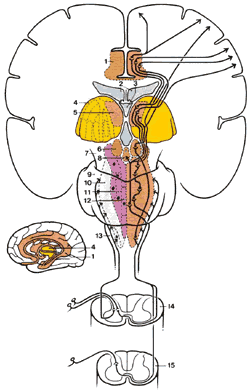 |
Рис.2.7. Диаграмма показывает пути вдоль латерального спинно-ретикулярного тракта из спинного в головной мозг. 1.Лимбическая структура переднего мозга 2.Латеральные желудочки 3.Третий желудочек 4.Таламус 5.Медиальное интраламинарное таламическое ядро 6.Гипоталамус 7.Средний мозг 8.Околоводопроводное серое вещество (ОСВ) 9.Варолиев мост 10.Латеральная зона ретикулярной формации 11.Медиальная зона ретикулярной формации 12.Медианная зона ретикулярной формации 13.Продолговатый мозг 14.Цервикальный столб 15.Люмбарный столб |
Спинно-мезэнцефалический тракт (СМТ)
Волокна СМТ (рис.2.8) заканчиваются в сером околоводопроводном веществе (СОВ) и других мезэнцефалических ядрах. СМТ берет начало в слоях I и V и восходит как контрлатерально, так и ипсилатерально . Полагают, что некоторые из СМТ-нейронов, которые заканчиваются в таламусе и лимбической системе, имеют различительную (дифференцирующую), а также вегетативную и рефлексивную активность и мотивационные функции. В дополнении к этому, СМП через связь с СОВ может активировать нисходящие ингибиторные системы, которые продуцируют анальгезию.
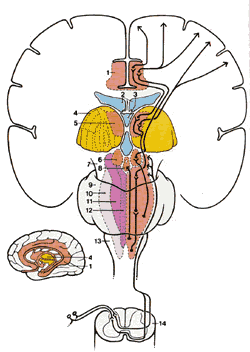 |
Рис.2.8. Диаграмма показывает пути вдоль латерального спинно-мезэнцефалического тракта из спинного в головной мозг. 1.Лимбическая структура переднего мозга 2.Латеральные желудочки 3.Третий желудочек 4.Таламус 5.Медиальное интраламинарное таламическое ядро 6.Гипоталамус 7.Средний мозг 8.Околоводопроводное серое вещество (ОСВ) 9.Варолиев мост 10.Латеральная зона ретикулярной формации 11.Медиальная зона ретикулярной формаций 12.Медианная зона ретикулярной формации 13.Продолговатый мозг 14. Люмбальный столб |
Спинно-цервикальный тракт (СЦТ)
СЦТ локализуется в дорсолатеральном канатике и восходит ипсилатерально, заканчиваясь в латеральном цервикальном ядре (рис.2.9). Затем волокна из латерального цервикального ядра проходят к контрлатеральному таламусу. Полагают, что так же, как и неоспинно-таламический путь, СЦП причастен к пространственному и временному различию болевых стимулов.
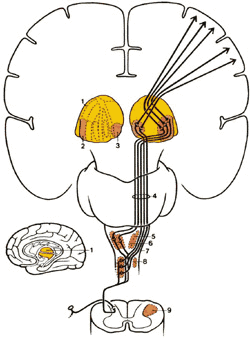 |
Рис.2.9. Диаграмма показывает пути вдоль латерального спинно-цервикального тракта из спинного в головной мозг. 1. Таламус 2.Nucleus ventralis posterior lateralis 3.Задняя группа таламических ядер 4.Медиальная петля 5. Nucleus cuneatus 6. Nucleus gratis 7.Каудальный продолговатый мозг 8.Спинно-цервикальный тракт |
Нисходящие механизмы болевого контроля
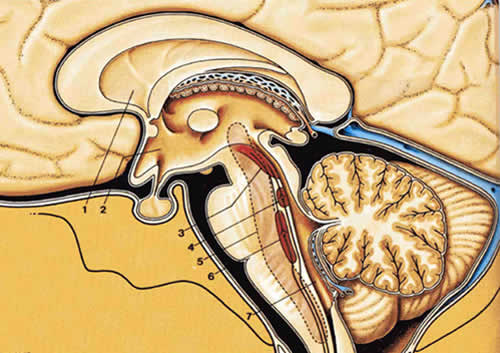
Стимуляция СОВ в среднем мозге как электрически, так и микроинъекциями морфина вызывает выраженную анальгезию. Проекции из СОВ на большое соединительное ядро и ядра ретикулярной формации мозгового ствола нисходят через дорсальный латеральный канатик к нейронам дорсального рога (рис.2.4).Полагается, что эти нисходящие волокна высвобождают 5НТ, которое подавляет активацию клеток желатинозной субстанции (рис.2.5). В дополнении к этому нисходящие волокна, которые берут начало в голубоватом месте, выделяют норэпинефрин, также подавляющий активацию клеток желатинозной субстанции. Считается, что антиноцицептивное действие адренэргических агентов ответственно за анальгетический эффект интраспинального клонидина.
| Рис.2.10. Структуры, вовлеченные в нисходящие механизмы контроля боли |
 |
|
1.Правый латеральный желудочек 2.Третий желудочек 3.Область околоводопроводного серого вещества 4.Голубоватое место 5.Ретикулярная формация 6.Nucleus raphe magnus 7.Четвертый желудочек |
Висцеральные болевые пути
Вегетативные афферентные нейроны, проходящие из висцеральных органов к стволу головного мозга и спинному мозгу, являются висцеральными афферентными компонентами спинных и краниальных нервов. Тела клеток этих нейронов так же, как таковые в соматических афферентных нейронах, локализованы в ганглии дорсального корешка соответствующих нервов (рис.2.11). Волокна висцеральных афферентных нервов проходят сквозь вегетативные ганглии, но не образуют в них никаких синаптических соединений, и входят в спинной мозг или ствол головного мозга в связке с соматическими афферентными волокнами тех же нервов. И висцеральные, и соматические афферентные нервные волокна образуют рефлексивные соединения с преганглионарными вегетативными нейронами в головном и спинном мозге. Следовательно, они функционально относятся к вегетативным нервам, но не являются их анатомическими компонентами.
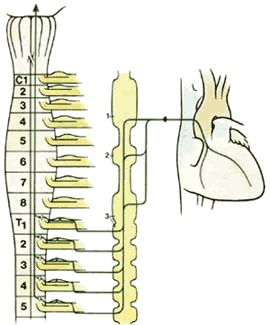 |
Рис.2.11. Схематичная диаграмма афферентных симпатических путей от сердца. Заметим, что как и все афферентные нервы, клетки тел лежат в пределах ганглиев дорсальных корешков. 1.Верхний цервикальный ганглий 2.Средний цервикальный ганглий 3.Сателлитный ганглий. |
Висцеральные афференты сходятся на нейронах дорсального рога так же, как и соматические ноцицептивные афференты. Повреждающие висцеральные стимулы передаются в головной мозг вместе с повреждающими соматическими стимулами посредством спинно-таламических путей. Считается, что висцеральная боль является результатом высокочастотного возбуждения висцеральных афферентов, которые обычно служат посредниками больше в гомеостатических, чем ноцицептивных функциях. Итак, висцеральные афференты могут возбуждаться на низких частотах в ответ на среднее расширение полого органа, но они возбуждаются на очень высоких частотах, когда имеет место чрезмерное растяжение. Эти высокие частоты активируют системы болевой проекции, начинающиеся в дорсальном роге. Результирующая боль, следовательно, воспринимается как происходящая из соматических областей, чьи нейроны проецируются на те же сегменты спинного мозга.
В таблице 2.3 перечислены внутренние органы и их сегментарная ноцицептивная нервная поддержка.
Таблица 2.3. Сегментарное распределение симпатических нервов
| Голова, шея, верхние конечности | Т1-5 |
| Сердце | Т1-5 |
| Легкие | Т2-4 |
| Пищевод | Т5-6 |
| Желудок | Т6-10 |
| Тонкий кишечник | Т9-10 |
| Толстый кишечник | Т11- L 2 |
| Печень и желчный пузырь | T7-9 |
| Поджелудочная железа и селезенка | T6-10 |
| Почки и мочеточник | T10- L 2 |
| Надпочечники | T 8- L 1 |
| Яички и яичники | T10-11 |
| Мочевой пузырь | T11- L 2 |
| Простата | T11- L 1 |
| Матка | T10- L 1 |
Необходимо хорошо различать иннервацию висцеральной структуры при принятии решения об уровне спинальной анестезии, который будет адекватным для хирургического вмешательства на этих структурах. Например, хотя кожная иннервация мошонки начинается с крестца, хирургия, проводимая на яичках, потребует спинальной анестезии, распространенной, по меньшей мере, до дерматома Т10, поскольку висцеральные афференты, ассоциированные с яичками, проходят с симпатическими волокнами, входя в центральную нервную систему на уровне Т10. В качестве другого примера рассмотрим кесарево сечение, которое требует среднегрудного уровня спинальной анестезии для уверенного обеспечения комфорта пациентки. Вероятно, это обусловлено тем фактом, что висцеральный и полосной перитонеум является огромной непрерывной структурой. Например, когда спинальная анестезия не распространена достаточно высоко, чтобы блокировать перитонеальные болевые волокна, входящие в спинной мозг в наиболее краниальной точке, результатом вытяжения (тракции) матки может стать перитонеальное раздражение и боль. Иногда, во время хирургическиъх вмешательств в абдоминальной области, несмотря на высокий уровень анестезии, блок остается неадекватным, поскольку пациент ощущает дискомфорт, тошноту и рвоту. Причиной является неудача в блокировании всех волокон, центрально передающих ощущения, включая блуждающий нерв.
Сердечно-сосудистые эффекты спинальной анестезии
Артериальная гипотония различной степени является обычным физиологическим свидетельством успешного спинального блока. В одном исследовании, которое включало более 11 000 случаев, гипотония наблюдалась у 38% пациентов. Основной причиной артериальной гипотонии во время спинальной анестезии является блокада волокон преганглионарного симпатического нерва (рис.1.31). Степень гипотонии связана с распространением агента местного анестетика в пределах субарахноидального пространства и расширением симпатической блокады (рис.2.12).
Рис.2.12. Сердечно-сосудистые изменения во время спинальной анестезии на различных сегментарных уровнях. Предполагается, что все пациенты положены горизонтально. ПС – периферическое сопротивление, ЧСС – частота сердечных сокращений, СВ – сердечный выброс, ЦВВ – центральный венозный возврат, C – сократимость, САД – среднее артериальное давление, НЕТ – изменения не наблюдались. | ||||||
| ПС | ЧСС | СВ | ЦВВ | С | САД | |
| НЕТ | НЕТ | НЕТ | НЕТ | НЕТ | НЕТ | |
| НЕТ | НЕТ | |||||
Например, низкий блок, ограниченный люмбальными и сакральными дерматомами, обуславливает небольшое изменения артериального давления или полное отсутствие таких изменений. Результатом высокой спинальной анестезии является симпатическая блокада волокон, иннервирующих сердце, а также контролирующих периферическое сосудистое русло и может, следовательно, вызвать выраженную гипотонию. Симпатическая блокада обычно распространяется на спинальные сегменты выше сенсорной блокады. Тогда как общепринято считать, что разница составляет 2 спинальных сегмента, новейшие исследования показывают большую вариабельность в этом отношении. Некоторые находят разницу в шесть сегментов, тогда как другие не показывают ни одного.
Физиологическая регуляция артериального давления
Артериальное давление определяется периферическим сопротивлением сосудов и сердечным выбросом (рис.2.13).
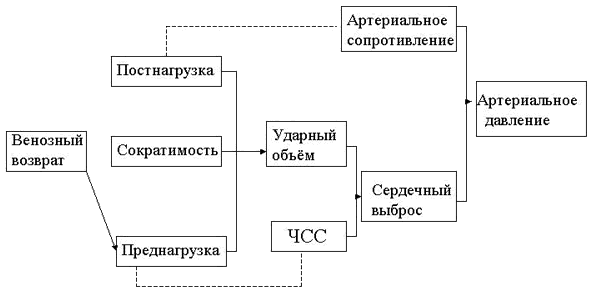 |
| Рис.2.13. Факторы, определяющие сердечную деятельность и артериальное давление. Представлено с разрешения Mark и Steel. |
Сердечный выброс это функция частоты сердечных сокращений и ударного объема. Ударный объем определяется внутренней сократимостью сердечной мышцы, сопротивлением, которое необходимо преодолеть сердцу (постнагрузка) и давлением наполнения в левом желудочке (преднагрузка), которое зависит от венозного возврата. Таким образом, изменения в артериальном давлении непосредственно связаны с изменениями или сосудистого сопротивления, или сердечного выброса, или и того и другого вместе. На сердечный выброс в свою очередь влияет изменения в преднагрузке, постнагрузке, сократимости миокарда и частоте сердечных сокращений. Периферическое сопротивление сосудов контролируется симпатической иннервацией артериол и локальной метаболической средой (рН ткани и крови, концентрациями СО2 и О2 ). Таким образом, артериальная гипотония во время спинальной анестезии связана с распространением симпатической блокады, которая влияет на периферическое сопротивление сосудов и сердечный выброс. Следовательно, величина сердечно-сосудистых изменений зависит от степени симпатического тонуса, который имеет место непосредственно перед спинальной инъекцией.
Периферическое сосудистое сопротивление
Симпатическая блокада, ограниченная нижней грудной и среднегрудной областью, приводит к вазодилятации в нижних конечностях с компенсаторной вазоконстрикцией в верхних конечностях. Таким образом, периферическое сосудистое сопротивление только слегка уменьшается, ограничивая степень гипотонии. По мере расширения блока вверх вазодилятация увеличивается, в то время как возможность компенсировать ее вазоконстрикцией уменьшается.
Сердечный выброс
Спинальная анестезия, которая распространяется только до среднегрудного уровня, не обуславливает значимых изменений в сердечном выбросе, если она проводится у пациента, находящегося в положении лежа или с опущенным головным концом (рис.2.12).
Однако, спинальная анестезия, распространяющаяся до верхнего грудного и шейного уровня, может стать причиной снижения сердечного выброса, обусловленного изменениями частоты сердечных сокращений, венозного возврата и сократимости миокарда.
Частота сердечных сокращений
Симпатические волокна, выходящие из Т1-Т5, контролируют частоту сердечных сокращений. Спинальная анестезия, которая блокирует эти волокна, обуславливает значимую сердечную симпатическую денервацию (рис.2.12). Поскольку в норме, присутствует симпатическая иннервация сердца, эта денервация является причиной умеренного снижения частоты сердечных сокращений. Выраженная брадикардия, наблюдаемая у некоторых пациентов во время высокой спинальной анестезии возможно, объясняется увеличенной вагальной активностью, которая встречается во время вазовагального синкопа или интракардиальных рефлексов.
Ударный объем
Ударный объем может уменьшаться во время высокой спинальной анестезии вследствие снижения венозного возврата и уменьшения сердечной сократимости (рис.2.12).
Венозный возврат
Венозный возврат у пациентов, перенесших симпатэктомию, очень сильно зависит от гравитации и, следовательно, положения. Симпатический контроль венозной системы является основным в поддержании венозного возврата и, следовательно, сердечно-сосудистого гомеостаза во время постуральных изменений. Вены формируют систему низкого давления и содержат наибольшую часть всего объема циркулирующей крови (приблизительно 70%). Когда спинальная анестезия вызывает симпатическую блокаду, этот контроль утрачивается, и венозный возврат становится полностью зависимым от гравитации. Если нижние конечности находятся ниже правого предсердия, денервированные вены будут расширяться, секвестрируя огромный объем крови. Возникающее в результате снижение венозного возврата, а, следовательно, и сердечного выброса, в сочетании с уменьшением периферического сопротивления могут стать причиной серьезной гипотонии.
Сократимость
Блокада симпатической иннервации сердца может стать причиной уменьшения инотропизма, приводящего к небольшому (около 15%) снижению сердечного выброса.
Синкопе
Клиницисты, имеющие опыт в проведении спинальной анестезии, распознают клиническую картину быстро развивающейся гипотонии, обычно сопровождающейся тошнотой и иногда рвотой. Могут иметь место кратковременная (моментальная) остановка сердца и потеря сознания, а также потоотделение и бледность во время восстановления. Это бывает только у пациентов, находящихся в сознании, и никогда не наблюдается у пациентов, которым дается сопутствующая общая анестезия. Сходство с вазовагальным обмороком является поразительным и блуждающий нерв, возможно, играет свою роль в этом случае.
Главной особенностью синкопе является внезапное падение венозного возврата, приводящее к значительному снижению сердечного выброса. Нормальным ответом на это является тахикардия и вазоконстрикция, призванные предотвратить венозную дилатацию.
Во время синкопе имеет место брадикардия, обусловленная быстрым снижением сердечного выброса. К счастью симпатическая активность восстанавливается и при своевременном устранении помех для венозного возврата (напр. таких, как поднятый вверх головной конец или IVC окклюзия), артериальное давление и частота сердечных сокращений увеличиваются. Если имеет место потеря сознания, восстановление часто сопровождается коротким эпилептиформным припадком. Это может быть спутано с токсической реакцией, хотя токсическое воздействие практически невозможно при малых дозах раствора местного анестетика, применяемых для спинальной анестезии.
Роль блуждающего нерва во время синкопе неясна. Синкопе, наблюдаемое у волонтеров после введения вазодилятаторов, все-таки имеет место, несмотря на тахикардию, обусловленную большими дозами атропина, введенного внутривенно. Было установлено, что интракардиальные рефлексы, которые предотвращают избыточную сократимость в не полностью заполненных желудочках, снижают как частоту сердечных сокращений, так и силу сокращения, когда венозный возврат радикально снижается. Полагают, что эти механизмы призваны предупредить механическое повреждение миокарда желудочка и снизить миокардиальное потребление кислорода.
Какими бы ни были механизмы, вовлеченные в синкопе, клиницист должен понимать, что нарушен именно венозный возврат и именно он подлежит восстановлению. Синкопе намного чаще встречается при поздних беременностях, чем у небеременных пациенток, поскольку IVC окклюзия может все-таки иметь место, несмотря на латеральный наклон или смещение матки. На ранней стадии развития гипотонии должен быть введен симпатический вазоконстриктор, предпочтительно с альфа- и бета-активностью, чтобы предотвратить атаку синкопе. С целью улучшения венозного возврата также следует быстро изменить положение пациента и, хотя кажется логичным использовать для лечения брадикардии атропин, от этого следует воздержаться, так как атропин не увеличивает венозный возврат и не влияет на артериальное давление во время синкопе. Сэр Томас Льюис, первым описавший вазовагальное синкопе и его лечение, указывал, что симпатический препарат, такой как эфедрин, обеспечивает как вазоконстрикцию, так и тахикардию, необходимые для быстрого восстановления.
Итак, причины гипотонии во время спинальной анестезии связаны с изменениями периферического сопротивления сосудов и сердечного выброса, которые зависят от изменения венозного возврата (рис.2.12). У большинства пациентов степень артериальной гипотонии обычно клинически не существенна, если венозный возврат поддерживается около нормального значения.
Факторы, влияющие на артериальную гипотонию, вызванную спинальной анестезией
Уровень симпатической блокады
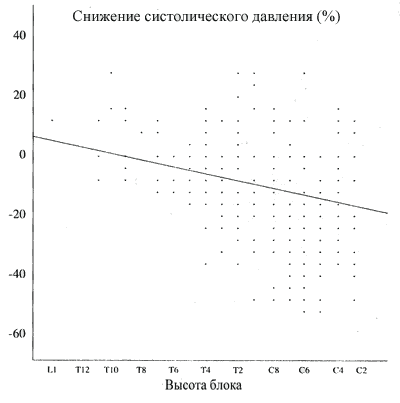
В то время как более высокий блок ассоциируется с более высокой частотой гипотонии, невозможно предсказать степень гипотонии у конкретного пациента на основе уровня спинальной анестезии. Например, у некоторых пациентов гипотония развивается при нижних грудных уровнях анестезии, тогда как у других с уровнями анестезии в верхней грудной и нижней шейной области гипотония незначительна или совсем отсутствует (рис.2.14). Эти индивидуальные различия отражают различия во взаимоотношениях между уровнем симпатической блокады и уровнем сенсорной анестезии, а также степенью симпатического тонуса, имевшего место перед началом спинальной анестезии.
| Рис.2.14. Взаимосвязь между процентным изменением систолического давления и высотой блока у 148 пациентов, получавших спинальную анестезию ( D . B . Scott , неопубликованные данные). |
 |
Положение пациента
На гипотонию во время спинальной анестезии существенным образом влияет положение пациента (рис.2.15).
| Рис.2.15. Диаграмма, показывающая влияние уровня блока и положения пациента на изменения различных сердечно-сосудистых показателей. ПС –периферическое сопротивление, СВ – сердечный выброс, САД – среднее артериальное давление, НЕТ – изменения не наблюдались. | |||
| ПС | СВ | САД | |
| НЕТ | НЕТ | НЕТ | |
| НЕТ | |||
У пациентов, помещенных с приподнятым головным концом, во время спинальной анестезии может развиться тяжелая гипотония, обусловленная венозным застоем. Следовательно, пациенты во время проведения спинальной анестезии должны находиться в положении со слегка опущенным головным концом, так как это способствует поддержанию нормального венозного возврата.
Факторы, связанные с состоянием пациента
Степень гипотонии, развивающаяся при одном и том же уровне спинальной анестезии может варьировать в зависимости от физического состояния пациента, что связано с базальным симпатическим тонусом.
Гиповолемия может стать причиной серьезного угнетения сердечно-сосудистой системы во время спинальной анестезии. При гиповолемии давление крови поддерживается повышенным симпатическим тонусом, который обуславливает периферическую вазоконстрикцию. Симпатический блок, являющийся следствием спинальной анестезии, может привести к выраженной гипотонии. Следовательно, гиповолемия является относительным противопоказанием к спинальной анестезии. Однако, спинальная анестезия вполне приемлема, если при наличии гиповолемии нормоволемия может быть достигнута замещением объема.
Беременные особенно чувствительны к симпатической блокаде и гипотонии, связанными со спинальной анестезией. Это происходит из-за механической обструкции венозного возврата беременной маткой. Беременные должны быть помещены в положение со слегка опущенным головным концом (латеральную наклонную позицию) немедленно после вводной части спинальной анестезии, чтобы предотвратить аорто-кавальную компрессию. Нарушения кровообращения, связанные со спинальной анестезией у беременных обсуждаются более детально в следующих публикациях.
У пожилых пациентов, страдающих гипертензией и ишемической болезнью сердца, гипотония во время спинальной анестезии развивается чаще по сравнению с молодыми здоровыми пациентами.
Факторы, связанные с анестетиками
Степень гипотонии может зависеть от препарата, которым выполняется спинальная анестезия. Несмотря на достижение одинакового уровня сенсорной анестезии, бупивакаин вызывает меньше гипотоний, чем тетракаин. Разница в гипотонии может быть связана с большей симпатической блокадой В-волокон при применении тетракаина (как было продемонстрировано в исследованиях in vitro ) в сравнении с бупивакаином.
Баричность анестезиологического раствора также может влиять на гипотонию во время спинальной анестезии. Гипербарические растворы тетракаина и бупивакаина приводят к большей гипотонии, чем изобарические растворы. Эти различия, вероятно, связаны с различиями в уровнях сенсорного и симпатического блока. Гипербарические растворы распространяются дальше, чем изобарические или гипобарические растворы и обуславливают большую симпатическую блокаду.
Сопутствующая (вспомогательная) общая анестезия предположительно увеличивает частоту гипотоний во время спинальной анестезии. Известно, что это же происходит в случаях сочетания общей анестезии с эпидуральным блоком (более детально об этом – в следующих публикациях).
Суммируя сказанное, отметим, что изменения в сердечно-сосудистой системе во время спинальной анестезии являются нормальным физиологическим ответом и обусловлены блокадой симпатических волокон. Знание факторов, ответственных за изменение артериального давления позволяет избежать тяжелой гипотонии. Такое знание также значимо при выборе подходящей терапии для восстановления угнетенных функций сердечно-сосудистой системы.
Респираторные эффекты спинальной анестезии
Влияние спинальной анестезии на функцию дыхания связано с уровнем блокады проводимости. Распространение спинальной анестезии до среднегрудного уровня или ниже редко обуславливает значимые изменения легочных функций. Однако, спинальная анестезия, которая распространяется до верхнегрудных дерматомов может стать причиной нежелательных изменений функции дыхания (напр., у пациентов с хроническими обструктивными заболеваниями дыхательных путей). С другой стороны у пациентов, страдающих ожирением, может наступать улучшение оксигенации, предположительно за счет увеличения растяжимости грудной клетки, которое в свою очередь является следствием паралича абдоминальных мышц. Среднегрудная спинальная анестезия не имеет значительного влияния на дыхательный объем, частоту дыхания, минутную вентиляцию или Р CO 2 в конце выдоха. Однако остановка дыхания может случиться вследствие непреднамеренной тотальной спинальной анестезии в связи с параличом дыхательных мышц и/или ишемией головного мозга, развившейся вследствие тяжелой гипотонии.
Контроль дыхания
Исследование, включавшее пациентов, которым без премедикации проводилась спинальная анестезия изобарическим бупивакаином и гипербарическим тетракаином, показало снижение остаточного РСО2 в конце выдоха примерно на 0.3-0.7 kPa . Повторное вдыхание СО2 увеличивало минутную вентиляцию и окклюзионное давление во рту. Ответ на СО2 во время спинальной анестезии обратно коррелирован с возрастом, напр., у молодых пациентов минутная вентиляция увеличивается значительно больше, чем у пожилых.
Спинальная анестезия, распространенная до середины верхних грудных отделов иногда вызывает диспноэ, которое не обусловлено слабостью дыхательных мышц. Диспноэ может происходить из дыхательных центров головного мозга, которые во время спинальной анестезии лишены афферентного ввода (сигналов) от абдоменальной области и грудной клетки. Стимул к дыханию в норме ингибируется этими проприоцептивными импульсами. Когда они заблокированы высокой спинальной анестезией, имеет место возросший стимул к дыханию, обусловленный снижением остаточного РСО2 в конце выдоха и увеличением вентиляторной реактивности на СО2. Пациент может воспринимать эти увеличенные вентиляторные стимулы как диспноэ.
Респираторные механизмы
Спинальная анестезия значимо не влияет на механизмы спокойного дыхания. Функциональная остаточная емкость снижается при минимальном изменении дыхательного объема. Измерения на вдохе давления, потока и объема указывают лишь на небольшое ухудшение функций инспираторных мышц даже при высоких грудных уровнях анестезии. Однако имеется взаимосвязь между уровнем спинальной анестезии и активностью экспираторных мышц. Высокие уровни спинальной анестезии приводят к заметному снижению максимального давления и скорости потока на выдохе. Снижение пиковой объемной скорости непосредственно зависит от направления распространения анестезии, которое получается выше, когда применяются гипербарические растворы. Влияние спинальной анестезии на функции мышц вдоха и выдоха связаны с тем, что при вдохе и выдохе вовлекаются различные мышцы. Главной мышцей вдоха является диафрагма. Другие мышцы вдоха включают внешние межреберные и грудино-ключично-сосцевидные мышцы. Диафрагма иннервируется спинномозговыми нервами С3-5. Таким образом, даже высокие грудные уровни не подавляют сжатие диафрагмы. В действительности функция диафрагмы может быть улучшена во время спинальной анестезии, поскольку сила, которую диафрагма должна развить, чтобы сместить внутренние органы абдоминальной области во время вдоха, уменьшается из-за паралича абдоминальных мышц. Диафрагма, следовательно, компенсирует паралич внешних межреберных мышц. На добавочные мышцы вдоха оказывает влияние лишь спинальная анестезия, достигающая верхних шейных нервов.
У здорового человека при спокойном дыхании выдох пассивен, но во время гипервентиляции он становится активным. Главными экспираторыми мышцами являются абдоминальные мышцы. Внутренние межреберные мышцы также участвуют в выдохе. Активные экспираторные усилия необходимы для эффективного кашля. Пациенты с тяжелым хроническим легочным заболеванием могут вовлекать экспираторные мышцы во время спокойного дыхания из-за увеличенного сопротивление выдыхаемому воздушному потоку и снижения эластичного ответа легких. Следовательно, у этих пациентов может развиться дыхательная недостаточность во время спинальной анестезии.
Регионарная анестезия, включающая эпидуральную или спинальную анестезию, часто предлагается как предпочтительная по сравнению с общей анестезией для пациентов, страдающих астмой. Поскольку пациентам-астматикам требуются активные усилия на выдохе, высокие уровни спинальной анестезии могут теоретически ухудшить эту функцию. Однако, исследования пациентов с хроническими заболеваниями легких указывают, что изменения респираторных механизмов в связи со спинальной анестезией не превышают допустимых порогов. Например, спинальная анестезия не обуславливает значительного изменения форсированной жизненной емкости легких у пациентов, имеющих заболевания легких, ни в положении на спине, ни в литотомическом положении. В действительности форсированный объем выдоха даже слегка увеличивается в литотомическом положении. В противоположность этому у здоровых людей положение влияет на функцию выдоха. Например, форсированная жизненная емкость легких снижается на 10% в положении на спине и 15% в литотомическом положении, тогда как объем форсированного выдоха снижается на 6% и 7% в этих позициях. Улучшение форсированного объема выдоха во время спинальной анестезии у пациентов с легочными заболеваниями является свойством «вспомогательного возврата» диафрагмы, обусловленного весом внутренних органов абдоминальной области во время выдоха.
Газообмен в легких
Низкие уровни спинальной анестезии незначительно влияют на газообмен. Потребление кислорода и продукция углекислого газа снижаются на 10-20% вследствие спинальной анестезии. Однако, если спинальная анестезия распространяется не выше среднегрудных уровней, такие показатели как минутная вентиляция легких, дыхательное мертвое пространство, альвеолярно-артериальная разница кислорода или углекислого газа, внутрилегочный шунт или напряжение газа в артериальной крови изменяются незначительно или совсем не имеют значимых изменений.
Исследования пациентов с ожирением указывают на то, что значения газов крови при спинальной анестезии у них не изменяются или даже фактически улучшаются. Субарахноидальная блокада, распространяющаяся до Т4-10 у пациентов, на 47%превышающих идеальную массу тела, увеличивала среднее напряжение кислорода в артериальной крови примерно на 1 кРа, тогда как напряжение углекислого газа и рН артериальной крови не изменялись. Улучшение растяжимости грудной клетки вследствие паралича абдоминальных мышц во время спинальной анестезии считается ответственным за улучшение вентиляционно-перфузионного соотношения у таких пациентов с ожирением. В контрольной группе пациентов с нормальным весом никаких значимых изменений в значениях газов крови не наблюдалось.
Хотя минимальные изменения дыхательных функций имеют место при среднегрудных уровнях спинальной анестезии, необходимо помнить, что предоперационная и интраоперационная седация может привести к нарушениям легочного газообмена во время спинальной анестезии. Например, у некоторых пациентов пероральный прием темазепама приводит при спинальной анестезии к снижению насыщения кислородом артериальной крови. В добавление к этому, у многих пациентов, которым вводится внутривенная седация, как дополнение к спинальной анестезии, наблюдается десатурация. Таким образом, при вводе седативных препаратов по возможности всегда должен осуществляться мониторинг оксигенации с помощью пульсоксиметрии.